Is Life a Miracle or Inevitable?
yuval bloch
The possibility of life on other planets has fascinated humanity for centuries. To make sense of the silence, we have long relied on the Drake Equation—a mathematical framework for estimating the number of active, communicative civilizations in our galaxy.
Conventional wisdom suggests the universe should be teeming with life. With trillions of stars and countless “Goldilocks” planets, the math implies that technologically advanced societies should be common. This brings us to the Fermi Paradox: if the universe is a crowded party, why is it so quiet?
To find an answer, we can simplify the Drake Equation into three core pillars:
- $Ph$ (Physical): The number of suitable planets in the universe.
- $Bi$ (Biochemical): The probability of life actually emerging on one of those planets — the problem of abiogenesis.
- $Ev$ (Evolutionary): The probability of that life developing into an intelligent, technological civilization.
The simplified equation for the number of civilizations ($N$) becomes:
$$N = Ph \times Bi \times Ev$$
In recent decades, our data for $Ph$ has surged. We now estimate there are roughly $10^{22}$ stars in the observable universe, most of which host orbiting planets. For a long time, it seemed unlikely that the biochemical and evolutionary steps could be rare enough to negate so many opportunities.
But there is another paradox that tells a different story — one that does not ask why intelligent life is rare, but how life could have appeared at all. Even with $10^{22}$ stars and 13.8 billion years, life might still be a statistical miracle. It is called the Eigen’s Paradox.
Between Eigen’s Paradox and the Fermi Paradox
Life is complex. Anyone who has tried to balance a career and a social life knows this, but the complexity goes much deeper. Even the simplest primitive life we know contains a DNA molecule with more than 100,000 nucleotides arranged in a nearly perfect sequence. A single error in that sequence can prove fatal.
In a universe governed by **entropy—**the natural tendency toward disorder—life manages to defy expectations and create remarkable order.
Highly complex molecules like DNA cannot appear by pure chance. They arise through evolution. Assembly Theory explains that, instead of building structures randomly from scratch, life uses structures it has already built (those with high “fitness”) as templates for further complexity. To bootstrap this process, the very first life needed to achieve four things simultaneously:
- Reproduce: Use existing structures as building blocks for new ones.
- Conserve information: Ensure that “blueprints” are faithfully passed to the next generation.
- Modify information: Allow for slight changes — mutations — that enable adaptation.
- Perform function: Interact with the environment in ways that increase fitness.
Here lies the central catch-22. This system is already astonishingly complex. The first life could not have formed through biological evolution, because the mechanisms of evolution must exist before evolution can begin. That first step had to be achieved through pure chemistry.
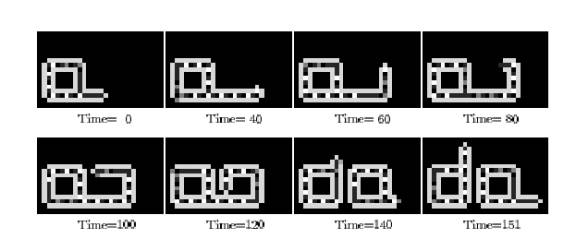
The Computational Model of Life
To appreciate how difficult this is, consider life as a computational entity. Replication is, at its core, a code-based task. Here is a simple example of a self-replicating program:
instructions = "print('instructions =',instructions); print(instructions)"
print('instructions =', instructions)
print(instructions)
This code prints itself. It requires two things: the instructions (the data) and a mechanism that executes them. Crucially, the instructions must encode at least two steps: copy the instructions, and build the mechanism that runs them. In biology, DNA plays the role of instructions, and enzymes serve as the
mechanism that executes them.
To estimate the minimal complexity life will require, ignoring any biochemical challenge. We can model them using Cellular Automata (CA)— grid-based systems in which cells update based on their neighbors. now let imigine a self replicating celolar aoutomata, in the simplest form the instruction often build as circle and the macnizhem is a simple “arm” geting out of it that can behive diffrently depend on the inforamtion it get from the circle, the arm moving around the circle and behvae diffrenly acording to the state of the inframtion cell it pass, now think how complex it is the arm need to be able to preform diffrent action depend on the state of the cell it meet, finish them and move to the next one, some of this action need to be copy the inforamtion it meet and some need to be follow them, the arm need a way to differ between the twom modes.
Current research suggests that if we want to both perform computational actions (which here represent metabolism) and reproduce, it might require around 150 cells. It is worth mentioning that only storing and reproducing information is much simpler. Think of it that adding more actions to an entity doesn’t sum up its complexity; it multiplies it as it needs to be able to differentiate between different types of instructions. And the more complex the mechanism is, the harder it becomes to copy
The Universal Lottery
Why do these small numbers matter? Because if the first replicator appeared through random chemical collisions, the math becomes terrifying.
- There are roughly $10^{22}$ planets in the observable universe.
- Each planet contains roughly $10^{50}$ atoms.
- The universe is approximately $10^{17}$ seconds old.
If every atom on every planet tried a new molecular combination every second, the universe has conducted roughly $10^{89}$ experiments since the Big Bang. If the simplest possible life form requires a specific arrangement of just 100 atoms, and we assume each of those positions can be filled by one of 10 different atom types chosen at random, the probability of assembling the right sequence by chance is roughly 1 in $10^{100}$.
It would never happen. Even in a universe full of planets, life would be a statistical impossibility—a true statistical miracle.
But before you start shopping for a place of worship, two factors fundamentally change the calculation:
- Life as a spectrum: The boundary between “non-life” and “life” is not a wall — it is a gradient. Chemistry is not purely random; its own rules bias the odds in favor of complexity.
- The RNA World: Early life may have functioned quite differently, using a single molecule that acted as both an instruction set and hardware simultaneously, and using the environment to bypass some calculations.
Life as a Spectrum
For life to reach its current complexity, all three conditions defined earlier must eventually be met. But satisfying even one of them is enough to begin accumulating complexity and edging toward life.
Consider autocatalytic molecules — chemical networks, such as certain sugars, that accelerate their own production. An existing sugar molecule increases the probability that more sugar molecules will form. A chain of sugar molecules does not replicate in the biological sense, meaning the information about how the chain evolved does not persist. But this simple self-amplification makes the complex building blocks of life far more common than pure chance would predict.
A second factor is the environment itself, which computational models tend to ignore. The leading theory for the origin of life holds that the first living organism was a self-replicating RNA molecule, and that self-replication was driven in two steps by the environment: when temperatures cool, new nucleotides attach to the existing RNA strand, extending it; when temperatures rise, the newly built strand separates from the template. The environment serves as an external clock and energy source, removing the need for the organism to encode those functions internally.
A Quiet Tiny Step, a Huge Leap for Science
This theory recently received striking experimental support. Scientists successfully created an information-preserving, self-replicating RNA strand from just 45 nucleotides — they named it qt45, short for Quite Tiny, 45. A nucleotide is a complex molecule, but it is also stable and produced by autocatalytic processes, so it would be expected to be abundant under the right conditions.
While qt45 can replicate and interact with its environment, it is not yet a fully functional candidate for Darwinian evolution, as it cannot perform any metabolic function. Without a function that can be improved by natural selection, evolution tends to favor simpler, faster-replicating variants — driving complexity downward rather than upward. But the gap between Qt45 and a true first ancestor of life is smaller than ever before.
What Is Still Missing
qt45 provides a plausible pathway to the origin of life, but the complete chain of events has not yet been fully filled in. A deeper understanding of prebiotic chemical reaction networks will tell us how probable the spontaneous appearance of nucleotides really is. Engineering more complex self-replicating RNA molecules that can also perform useful functions will reveal how difficult the jump from nucleotide to life truly was. And a more quantitative theory of evolution will tell us how probable — or improbable — the transition from simple to complex life actually is.
To know whether life exists beyond Earth, we may first need to understand far more carefully how it arose here.
But in the end, it may not matter whether life is a perfect statistical miracle or an abundance spread across the cosmos, or somewhere in the middle. Life is a wonder either way — something beautiful and intricate, so messy yet so organized. So let us not take it for granted.